Под названием витамин D объединяют группу родственных соединений, обладающих противорахитным эффектом. Важнейшие из них — эргокальциферол (витамин D2) и холекальциферол (витамин D3).
Кальциферолы образуются вследствие фотоизомеризации из природных стеринов (провитаминов D) при солнечной радиации или искусственном УФ-облучении. Провитамином эргокальциферола служит стерин растительного происхождения — эргостерин, провитамином холекальциферола — 7-дегидрохолестерин, присутствующий в коже и других тканях животных.
У цыплят очень богата провитамином кожа конечностей и бедра, кожа с участков, покрытых оперением; образованию эргокальциферола в коже способствует секрет копчиковой железы.
Интенсивность синтеза холекальциферола достаточно высока, но полностью не удовлетворяет потребности животных, в связи с чем они нуждаются в экзогенном поступлении витамина. Потребность дойных коров в холекальцифероле удовлетворяется за счет его эндогенного синтеза при содержании животных на пастбище или в открытых загонах 6—12 ч в сутки. При пасмурной погоде или кратковременной пастьбе синтез витамина недостаточен.
Особое внимание должно быть обращено на D-витаминную полноценность рационов при круглогодовом стойловом содержании коров и кормлении их брикетированными кормами, приготовленными из растений, предварительно не провяленных.
Необходимость регулярного и равномерного поступления витамина D в организм обусловливается тем, что резервы его в тканях невелики и весьма лабильны. Концентрация витамина D в плазме крови у разных видов животных составляет 1,5—3 мкг в 100 мл, редко достигая 5—6 мкг.
Для всех видов сельскохозяйственных животных физиологическая активность холекальциферола и эргокальциферола одинакова, за исключением птиц. У последних соотношение активностей этих соединений составляет 30:1—40:1, что обусловлено меньшим удержанием в организме дериватов эргокальциферола и их более низкой биологической эффективностью.
Отсутствие или резкий дефицит витамина D вызывает у взрослых животных остеомаляцию, а у молодняка рахит. Эти заболевания сопровождаются снижением произведения активностей ACa2+·AHPO42- в сыворотке крови и характерными биохимическими и гипопатологическими изменениями. Признаки заболевания проявляются у цыплят примерно через 2—3 недели, у лабораторных животных — через 4—5 недель, у поросят — через 5—6 недель после начала скармливания дефицитного по витамину D рациона.
Ранними симптомами недостаточности являются гипокальциемия, гиперфосфатемия и гипермагниемия. Однако в отличие от истинной низкокальциевой формы рахита при дефиците витамина D кальциемия может поддерживаться на постоянном уровне в результате активации функции паращитовидных желез. У кур-несушек при недостатке холекальциферола снижается (а затем прекращается) яйценоскость, уменьшается масса яиц, резко ухудшается качество скорлупы.
Известно, что у сельскохозяйственных животных даже прирезком избытке кальция в рационе, гиперкальциурия предупреждается либо быстрым поглощением абсорбируемого кальция скелетом (у растущих животных), либо гомеостатическим регулированием уровня абсорбции кальция, либо, наконец, увеличением его эндогенной кишечной экскреции. Иначе говоря, основной регулирующий механизм для кальция находится на уровне желудочно-кишечного канала.
Главным компонентом этого механизма является витамин D, стимулирующий абсорбцию кальция в кишечнике, что, в свою очередь, способствует повышению уровня кальция и фосфора в сыворотке крови и отложению минерального компонента кости. Другими предполагаемыми функциями витамина D являются: стимуляция отложения костного минерала; влияние на резорбцию кости; повышение реабсорбции фосфатов почками и снижение их выделения с мочой.
Механизм основного действия витамина D — его влияния на абсорбцию кальция — также трактуется по-разному. Если отбросить гипотезы, не выдержавшие испытания временем, то обсуждению подлежат две основные точки зрения:
- витамин D (или его дериваты) повышают способность ионов кальция к диффузии через стенку кишечника, противодействуя влиянию факторов, снижающих концентрацию Са2+, или увеличивая проницаемость мембраны кишечного эпителия;
- витамин D необходим для формирования или запуска специальной транспортной системы для кальция в кишечной стенке.
Обе точки зрения за последние годы обогатились новыми экспериментальными доказательствами, хотя полной ясности в вопрос об интимном механизме действия витамина D они не внесли.
При помощи меченного тритием витамина D удалось проследить за образованием и обменом его метаболитов в организме рахитичных животных. Основными метаболитами, циркулирующими в крови, оказались 25-оксихолекальциферол и 25-оксиэргокальциферол, обладающие в 1,5 раза большей активностью, чем исходный витамин.
Образуются в печени вследствие гидроксилирования витамина D в положении 25.
В кишечнике же активной формой витамина является 1,25-дигидроксихолекальциферол. Последний образуется в почках из 25-оксипроизводных витамина D в 4—5 раза более активен для предупреждения рахита, чем нативный витамин D.
В процессе метаболизма образуются и другие активные формы витамина, например сульфатированные.
Эта фракция составляет основную часть D-витаминной активности молока.
Значительная часть введенного витамина (от 30 до 70%) подвергается инактивации и выведению из организма разными путями (с желчью, калом, мочой, частично с молоком). В связи с этим желчь нельзя рассматривать как секрет, способствующий абсорбции кальция благодаря высокому содержанию в нем витамина D. При выключении желчных протоков всасывание кальция снижается как у нормальных, так и у рахитичных цыплят.
Образование в организме физиологически активных форм витамина D объясняется наличием 10—16-часового лаг-периода (периода между моментом перорального или внутривенного введения витамина в организм и началом его стимулирующего эффекта на абсорбцию кальция), а также отсутствие влияния витамина на абсорбцию в опытах на изолированных участках кишечника in vitro.
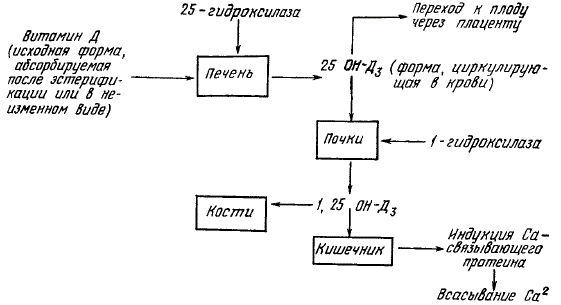
Схема метаболизма витамина D
Другим важным открытием было выделение из слизистой кишечника витамин D-зависимого низкомолекулярного водорастворимого кальцийсвязывающего белка.
Белок образует комплексы и с другими щелочноземельными элементами. Сродство белка к катионам убывает в следующем порядке: Ca, Sr, Ba, Mg.
Установлено, что его содержание в слизистой тесно коррелирует с увеличением абсорбции кальция и (до известного предела) с D-витаминной полноценностью рациона. У кур-несушек количество белка в слизистой кишечника увеличивается после снесения первого яйца и остается высоким в течение всего периода яйцекладки.
Высказано предположение, что 1,25-дигидрооксикальциферол индуцирует образование кальцийсвязывающего белка в цитоплазматической фракции эпителиальных клеток. Схематично это выглядит так. Специальные рецепторы на хромосомах ядер эпителия фиксируют 1,25-дигидрооксихолекальциферол. Последний оказывает депрессорный эффект, стимулируя биосинтез информационной РНК для кальцийсвязывающего белка. Информационная РНК несет соответствующую информацию в рибосомы цитоплазмы, где с помощью транспортной РНК происходит биосинтез белка. Антибиотик актиномицин D и синтетический аналог глюкокортикоидов преднизолон, введенные рахитичным животным перед дачей витамина D, ингибируют стимуляцию витамином D абсорбции кальция в кишечнике и биосинтез кальцийсвязывающего белка. Поскольку актиномицин блокирует ДНК-зависимый синтез. РНК, это рассматривается как доказательство участия витамина D в синтезе РНК, кодирующих образование кальцийсвязывающего белка.
По мнению Вассермана, аналогичным образом при участии производных витамина D осуществляется синтез кишечной АТФ-азы и щелочной фосфатазы — ферментов, играющих важную роль в механизме всасывания ионов кальция и фосфата.
Эта довольно стройная теория имеет, однако, уязвимые места. Возражения оппонентов сводятся к следующему:
а) не подтверждены убедительно данные о связывании меченого витамина D хроматином ядер; локализацию же его на мембране ядра можно рассматривать как облегчение доступа к ядерному материалу именно ионов Са2+, а не витамина;
б) увеличение синтеза РНК в эпителии кишечника у рахитичных животных под влиянием витамина D свидетельствует лишь об общем усилении ДНК-зависимого синтеза РНК и белков вследствие ускоренного роста слизистой. Соответственно следует оценивать и изменение микроструктуры клеточного эпителия под влиянием витамина D;
в) подавление биохимического эффекта витамина D актиномицином и другими ингибиторами не является доказательством роли витамина D как генетического индуктора. Его можно интерпретировать как отражение поврежденного общего синтеза нуклеиновых кислот и белков в быстрообновляющихся клетках слизистой кишечника;
г) актиномицин нарушает процесс превращения 25-оксихолекальциферола в 1,25-диоксихолекальциферол. Введение последнего рахитичным животным вызывает быстрое и значительное усиление всасывания кальция в тонком кишечнике, которое не снимается актиномицином;
д) в двенадцатиперстной кишке витамин D усиливает перенос ионов Са2+ в обоих направлениях, т. е. механизм его действия связан с увеличением проницаемости стенки кишечника. Усиление диффузии Са2+ через стенку под действием витамина D может быть отчасти объяснено его подавляющим влиянием на поглощение кальция митохондриями.
Таким образом, можно констатировать, что основной механизм действия витамина D на абсорбцию кальция остается пока недостаточно ясным.
Очевидно, что в организме витамин D подвергается превращениям, в процессе которых образуются высокоактивные дериваты, по-видимому, влияющие на разные стороны обмена. На уровне кишечника таким соединением является 1,25-диоксихолекальциферол, усиливающий образование кальцийсвязывающего белка. Однако роль 1,25-диоксихолекальциферола как генетического индуктора специфического белкового синтеза не доказана. Более правдоподобно предположение об усилении образования кальцийсвязывающего белка из неактивного предшественника, что связано с индукцией синтеза РНК и белков.
Не ясен также механизм участия специфического белка в переносе кальция. Возможно, что этот белок наряду с другими (синтез которых также зависит от витамина D) является компонентом насоса, осуществляющего активный перенос кальция против концентрационного и электрохимического градиента. Может, быть, связывая ионы кальция, а затем их освобождая, белок способствует созданию высокого концентрационного градиента на поверхности микроворсинок. Наконец, не опровергнута и гипотеза о влиянии витамина D на процессы пассивной и «облегченной» диффузии, которые также играют важную роль в абсорбции кальция.
Лучшее всасывание неорганического фосфата в кишечнике под влиянием витамина D обычно рассматривают как вторичный эффект, связанный с абсорбцией кальция. Однако имеются данные, что всасывание кальция и фосфатов это два независимых процесса, регулируемых витамином D. Основным местом всасывания кальция является двенадцатиперстная кишка, а фосфата — верхняя часть тощей кишки. Активные метаболиты, влияющие на абсорбцию фосфата и механизм их действия, неизвестны.
Влияние витамина D на гомеостаз фосфата в крови может быть прямым и косвенным. Прямое влияние связано с усиленным всасыванием фосфата и с повышенной реабсорбцией фосфатов в почках. Косвенное влияние обусловлено реципрокным снижением уровня паратгормона (при увеличении содержания кальция и фосфата в крови), что уменьшает экскрецию фосфата с мочой.
Неясным остается вопрос о влиянии витамина D на отложение или резорбцию кальция в скелете. В свете современных данных можно считать, что витамин D, по-видимому, влияет на процесс образования органического матрикса кости или его подготовку к кальцификации, но не на сам процесс минерализации.
Установлено, что витамин D стимулирует созревание эпифизарного хряща. Под воздействием витамина в хряще усиливается активность щелочной фосфатазы, гексозаминсинтетазы и пирофосфатазы. Увеличение уровня галактозаминов свидетельствует о влиянии витамина D на метаболизм хондроитинсульфата.
Что касается резорбции костной ткани, то этот процесс является сферой взаимодействия паратгормона и витамина D. В отсутствие витамина костные резервы под влиянием паратиреоидного гормона не мобилизуются. Даже ничтожная доза витамина D восстанавливает чувствительность костной ткани к гормону. В очень больших дозах витамин D имитирует эффект самого паратгормона, мобилизуя кальций из костей и усиливая экскрецию фосфатов почками. Возможно отчасти этим обусловлен положительный эффект ударных доз витамина D при некоторых формах нарушения половой цикличности у коров. Влияние гипокальциемического гормона тиреокальцитонина тоже, по-видимому, зависит от витамина D.
Приведенные выше данные свидетельствуют о наличии в организме комплексной и гибкой системы гуморальной регуляции метаболизма кальция и фосфора. Несомненно, однако, что наряду с гуморальными механизмами в регуляции принимают участие и нервные механизмы как центральные, так и периферические. Информация по этому вопросу пока только накапливается; имеющиеся данные обобщены в одной из работ.