Исследованию влияния ионизирующих агентов на мутационный процесс уделяется огромное внимание.
Это определяется тем значением, которое ионизирующие излучения приобрели в жизни человека в последние десятилетия. Однако далеко не все представляют себе, сколь тяжелы последствия повышения фона радиации. Это можно понять только на основе изучения закономерностей, влияния ионизирующих излучений на наследственную изменчивость. Известно, что облучение человека может привести его к гибели в силу лучевого поражения, но не менее опасными являются последствия облучения его половых клеток. Под влиянием даже незначительной дозы излучений резко возрастает частота мутаций. Подавляющее большинство мутаций порождает различные наследственные уродства и болезни. Накапливаясь в поколениях, они могут привести большие страдания человечеству. И обязанность современного общества состоит не только в том, чтобы сохранить жизнь ныне живущего поколения, но и оградить будущие поколения от наследственного отягощения вредными мутациями.
Вместе с тем ионизирующие излучения с большим успехом используются в селекции и медицине. Они широко применяются также для изучения многих биологических явлений.
Виды ионизирующих излучений
В современных генетических исследованиях широко используются различные ионизирующие излучения.
Мы должны иметь некоторое представление об их природе.
В настоящее время известно около 40 элементарных частиц. Однако доступными для биологических исследований являются лишь немногие, некоторые из них мы и рассмотрим.
1. Электрон — частица, обладающая массой покоя m0 = 9,06 х 10-28 г и единичным отрицательным электрическим зарядом е = 4,8X 10-10 ед. CGSE.
2. Позитрон. Масса покоя равна массе электрона. Обладает единичным положительным зарядом. Во всех взаимодействиях с веществом почта не отличается от электрона.
3. Фотон, иначе γ-квант, рентгеновский квант, световой квант. Обладает нулевой массой покоя и нулевым зарядом. Фотон представляет дискретную частицу электромагнитного излучения. Характеристикой его является энергия и связанная с ней частота электромагнитных колебаний.
4. Мезоны. Под этим названием объединяется более десятка элементарных частиц, обладающих весьма различными свойствами и имеющих массу, равную 200—1000 массам электрона.
5. Протон — частица с массой, равной 1834 электронным массам. Заряд равен единице, положительный.
6. Нейтрон — частица с массой покоя, равной 1836 электронным массам, и нулевым зарядом. Вместе с протонами входит в состав атомных ядер.
7. α-частицы представляют ядра атома гелия, состоящие из двух протонов и двух нейтронов. Они часто используются в биологических исследованиях.
Единицей измерения энергии перечисленных частиц является электрон-вольт, или энергия, которую приобретает частица с зарядом, равным заряду электрона при прохождении разности потенциалов в 1 вольт; 1 электрон-вольт (эв) = 1,6×1012 эрг; 1000 эв = 1 кэв, 1 000 000 эв = 1 Мэв.
Вещество построено из атомов, которые имеют ядро, состоящее из протонов и нейтронов, окруженное облаком (оболочкой) электронов. В результате возбуждения электронной оболочки возникает электромагнитное излучение различной энергии. При возбуждении внешних атомных электронов испускаются кванты видимого света и ультрафиолета. При возбуждении более глубоких оболочек возникают рентгеновские кванты, при возбуждении ядра атома — γ-кванты. Названия «рентгеновские» и «γ-кванты» являются весьма условными, так как обозначают лишь их происхождение.
Основными источниками рентгеновских квантов в эксперименте являются рентгеновские трубки, в которых используется эффект испускания ускоренными электронами электромагнитного излучения при торможении в электрическом поле ядра. Современные ускорители электронов (синхротроны и бетатроны) позволяют получать таким методом рентгеновские или γ-кванты с энергией до 200—300 Мэв.
Источником γ-квантов обычно являются возбужденные ядра атомов радиоактивных элементов, переходящие в основное состояние с испусканием γ-квантов. Примером является радиоактивный изотоп Со60, широко применяемый в различных исследованиях.
При радиоактивных превращениях также могут испускаться электроны или позитроны, обычно называемые в этом случае β- или или β+-частицами, и α-частицы. Основным источником быстрых тяжелых заряженных частиц являются ускорители, в которых протоны, дейтроны (ядра тяжелого водорода), α-частицы и ядра более тяжелых элементов, называемые многозарядными ионами, получают энергию до 1000 Мэв.
Источниками нейтронов обычно являются различные ядерные реакции, такие, например, как взаимодействие ускоренных дейтронов с ядрами тяжелого изотопа водорода трития, в результате которого получаются нейтроны с энергией около 14 Мэв. Наиболее мощным источником быстрых и медленных нейтронов являются ядерные реакторы, работающие на реакции деления урана.
Основным результатом воздействия ионизирующей частицы на атомы облучаемого вещества является ионизация — образование пар ионов, иначе говоря, высвобождение электронов из атомов или молекул, на что и затрачивается основная часть энергии падающей частицы, т. е. кванта. При этом мы сталкиваемся с тем удивительным фактом, что число пар ионов, образуемых в результате поглощения одинаковой энергии первичных ионизирующих частиц, не зависит от рода этих первичных частиц. Так, например, на образование пары ионов в воздухе или в живой ткани затрачивается 34 эв независимо от того, является ли первичная частица электродном, протоном или α-частицей. Это является следствием того, что ионизации, вызываемые первичной падающей частицей, сравнительно редки, а подавляющая доля ионизации происходит в результате вторичных процессов, ход которых мало зависит от природы первичного процесса взаимодействия. Таким образом, результаты воздействия разных ионизирующих излучений принципиально ничем не отличаются.
Однако, хотя конечный результат облучения одинаков для любых излучений, имеются существенные различия в характере ионизации, вызываемой различными первичными частицами. Эти различия заключаются в неодинаковой плотности ионизации, т. е. числе пар ионов, образующихся на единицу объема, и в неодинаковом распределении ионизации во всей толще облучаемого вещества.
Так как в конечном итоге любая частица выбивает атомные электроны, то прежде всего рассмотрим взаимодействие быстрых электронов с атомами вещества.
Ввиду того, что электрон обладает чрезвычайно малой массой покоя, его взаимодействие с ядрами атомов и атомными электронами существенно различно. При взаимодействии с ядром электроны испытывают рассеяние без изменения своей кинетической энергии. Это аналогично тому, как упругий шарик отскакивает от массивной плиты, изменяя при этом направление своего движения, но не меняя своей кинетической энергии. При взаимодействии с атомными электронами вследствие равенства масс обеих взаимодействующих частиц происходит полная или частичная передача энергии.
Оба этих процесса взаимодействия с ядром и атомным электроном примерно равновероятны, и поэтому путь электрона в веществе крайне извилист, и прохождение пучка электронов через слой вещества будет характеризоваться статистическими закономерностями. В силу этого ослабление пучка первичных электронов происходит постепенно.
Взаимодействие первичного электрона с атомными электронами приводит к образованию вторичных, быстрых электронов с распределением между ними всей или части энергии первичного электрона. Вторичные электроны в свою очередь как бы «размножаются» до тех пор, пока энергия образующихся вторичных частиц не станет слишком малой. Такой лавинообразный характер взаимодействия, приводящий, в конечном счете к возбуждению атомов и молекул вещества и образованию ионов, характерен для любого вида первичных частиц.
Вероятность взаимодействия первичного электрона с атомами зависит от его энергии. Чем больше энергия, тем меньше соударений испытывает первичный электрон в пересчете на единицу длины пробега.
Фотоны в основном взаимодействуют с электронной оболочкой атомов. При этом взаимодействие зависит от энергии фотона. Фотоны очень малой энергии, соответствующие видимому свету, могут возбуждать лишь внешние электронные оболочки атомов. Ультрафиолетовые лучи вызывают не только возбуждение электронных оболочек, но и образование так называемых фотоэлектронов. Этот процесс, называемый также фотоэффектом, заключается в том, что вся энергия фотона поглощается атомом и затем передается одному из электронов за вычетом энергии связи этого электрона в атоме.
Фотоэффект осуществляется со значительной вероятностью только в том случае, если энергия фотона не намного (не более чем в 2—3 раза) превосходит энергию связи электронов в атоме. С ростом энергии фотона вероятность фотоэффекта падает. При больших энергиях главную роль начинает играть комптон-эффект. Это процесс, в котором фотон непосредственно воздействует на атомный электрон, передавая ему часть своей энергии, в результате чего образуется так называемый электрон отдачи, или комптон-электрон. Фотон, потерявший часть своей энергии, может дальше еще раз испытать комптоновское рассеяние, и т. д. до полного поглощения.
Все перечисленные виды взаимодействия фотонов с атомам являются первичными. Результатом каждого из них будет образование быстрого электрона, поглощающегося затем в среде так же, как это описано для случая взаимодействия электронов с атомами.
Коэффициент поглощения для фотонов большой энергии очень мал, поэтому жесткие фотоны (γ-кванты) могут проходить без значительного ослабления большие толщи вещества. Для фотонов с энергией около 1 Мэв слой половинного поглощения будет равен примерно 26 см воды. Для электронов такой же энергии слой полного поглощения будет 0,5 см воды.
Рентгеновы лучи в зависимости от энергии квантов (величина которой и определяет характер их поглощения) разделяются на мягкие и жесткие.
Взаимодействие тяжелых заряженных частиц (протоны, α-частицы и многозарядные ионы) с атомами вещества имеет ряд особенностей, отличающих их от легких частиц (электронов и фотонов). Во-первых, благодаря своей большой массе тяжелые частицы при прохождении через толщу вещества мало изменяют свою траекторию. Во-вторых, в силу значительно меньшей скорости их движения (при энергии, равной энергии легких частиц) взаимодействие их с атомными электронами происходит значительно чаще. Однако энергия, передаваемая каждому электрону, сравнительно невелика. Потери энергии частицы на единицу пути значительно больше, чем в случае легких частиц.
Первое обстоятельство приводит к тому, что при данной энергии частицы (протона или α-частицы) пробег ее в веществе является строго определенным. Второе обстоятельство приводит к тому, что вся энергия частицы, расходующаяся на ионизацию, выделяется в очень малом объеме, прилегающем к ее траектории. Благодаря этому плотность ионизации очень велика, что приводит к сильному разрушению молекул, находящихся на пути частицы. Чем больше заряд частицы, тем выше создаваемая плотность ионизации.
Благодаря большим потерям энергии на единицу пути пробег тяжелых частиц, в веществе очень мал. Так, например, пробег протонов с энергией в 10 Мэв в воде будет составлять примерно 1,4мм. Для α-частицы с энергией 10 Мэв пробег в воде составляет всего лишь 0,15 мм. Глубина проникновения тяжелых частиц в вещество очень невелика. Поэтому естественно, что воздействие α-радиоактивных веществ (энергия а-частиц при этом 5—7 Мэв) ограничивается поверхностными тканями организма. Только в случае непосредственного введения радиоактивного вещества внутрь организма можно получить эффект воздействия α-частиц на внутренние ткани.
Взаимодействие нейтронов с веществом довольно сложно. Из-за отсутствия электрического заряда нейтроны могут взаимодействовать только с ядрами атомов. При этом нейтроны различных энергий обладают существенно различным воздействием. Нейтроны с энергией больше нескольких килоэлектронвольт при взаимодействии с ядрами атомов сообщают им часть своей энергии. Получающиеся ядра отдачи в дальнейшем взаимодействуют со средой, подобно тяжелым заряженным частицам, описанным выше. Именно они создают очень плотную ионизацию вблизи траектории. Особенно эффективно идет процесс образования ядер отдачи в водородсодержащих веществах, каковыми являются все живые ткани. В этом случае образуются протоны отдачи, обладающие энергией, в среднем равной половине энергии нейтронов.
Так как вероятность взаимодействия быстрых нейтронов с атомными ядрами очень мала, то нейтроны поглощаются веществом довольно слабо. Поэтому ядра отдачи и соответственно участки очень плотной ионизации, вызываемой ими, распределены по всей толще облучаемого вещества.
Нейтроны с очень малыми энергиями (единицы и доли электрон — вольта), называемые медленными, или тепловыми, поскольку их энергия близка к энергии теплового движения атомов и молекул, взаимодействуют с веществом главным образом путем образования γ-квантов, возникающих при захвате нейтрона ядром. Поэтому воздействие тепловых нейтронов на живую ткань сходно с воздействием γ-квантов.
Резюмируя вышеизложенное, можно сказать, что:
- воздействие легких частиц (электроны, фотоны) создает в веществе ионизацию малой плотности, рассеянную более или менее равномерно по всей толще облучаемого вещества,
- тяжелые заряженные частицы — протоны, α-частицы дают ионизацию с очень большой локальной плотностью,
- быстрые нейтроны вызывают аналогичное действие с той особенностью, что участки ионизации распределяются по всему облучаемому веществу,
- медленные (тепловые) нейтроны действуют аналогично легким частицам.
Образование ионов в облучаемой ткани не является конечным звеном воздействия ионизирующих излучений на живое вещество. Наоборот, оно является началом сложной цепи физико-химических процессов, развивающихся в течение некоторого времени и приводящих в конечном счете к перестройке молекул, изменению хода различных биохимических процессов, следствием которых могут быть мутации гена и разрывы хромосом.
Важным звеном этой цепи является радиационное разложение воды — основной составной части живых организмов. Показано, что ионизирующие излучения вызывают в воде образование перекиси водорода Н2O2 в измеримых количествах. Причем Н2O2 может образовываться как в присутствии кислорода, так и без него за счет возбуждения молекулы воды и соседних радикалов, например ОН + ОН* = Н2O2 или Н2O + Н2O* = Н2 + Н2O2 и т. д. (звездочкой отмечена возбужденная молекула).
Как мы увидим дальше, наличие или отсутствие кислорода в момент облучения оказывает большое влияние на частоту возникновения мутаций (кислородный эффект). Использование радиационной химии создает замечательные перспективы для познания процесса возникновения мутаций, химической и генетической дискретности хромосом.
В заключение приведем единицы измерения, которыми пользуются при определении степени облучения исследуемого вещества.
Доза ионизирующего излучения, поглощенного веществом, измеряется отношением поглощенной энергии (в эргах) к весу вещества (в граммах). Энергия излучения в 100 эрг, поглощенная 1 г облученного вещества, составляет 1 рад.
Дозу рентгеновых и γ-излучений принято измерять в рентгенах (р). Рентген — это доза излучения, при которой в см3 атмосферного воздуха при температуре 0° С и давлении 760 мм рт. ст. образуется около 2 млрд. пар ионов. При облучении воды и живой ткани дозой 1 р в 1 мк3 возникает около двух ионизаций. На образование одной пары ионов в воздухе или в живом веществе необходима эквивалентная энергия, равная 34 эв.
Мощность дозы рентгеновых и γ-излучений измеряется в рентгенах в 1 сек, в 1 мин и т. д.
В ряде случаев применяется единица дозы — биологический бивалент рентгена (БЭР). Эта единица определяется как доза облучения любого вида первичных частиц, дающая биологический эффект, такой же, как при облучении рентгеновскими или γ-квантов, дозой в 1 р.
Реакции клеток на облучение
Облучение клетки вызывает в ней глубокие изменения, проявляющиеся прежде всего в нарушении процессов деления.
Торможение клеточного деления ведёт к депрессии роста организма и поражению нормальной деятельности его тканей. Подавление митотической активности клетки при облучении объясняется действием ионизирующих излучений на ядро.
В облученной цитоплазме и в ее структурных элементах возникают различные изменения физико-химического характера: изменения вязкости, вакуолизация, повышение проницаемости для электролитов и т. д. Цитоплазма яиц морского ежа, тутового шелкопряда и других животных сохраняет жизнеспособность при облучении рентгеновыми лучами в дозах до нескольких десятков тысяч рентген, а в некоторых случаях — до сотен тысяч. Так, в опытах Б. Л. Астаурова яйца тутового шелкопряда облучались рентгеновыми лучами в дозе 550 000 р, которая у 100% яйцеклеток убивает ядро, но не разрушает цитоплазму. Если такие облученные яйцеклетки оплодотворить необлученными спермиями, то из яиц могут развиться андрогенетические особи. Однако чувствительность цитоплазмы к облучению у разных живых организмов разная. Так, в опытах с облучением яиц другого насекомого Habrobracon было показано, что развитие его андрогенетических особей тормозится уже при воздействии 15 000 р. В отличие от цитоплазмы ядро клетки поражается даже весьма малыми дозами облучения — порядка нескольких единиц рентгена. Различия в чувствительности ядра и цитоплазмы к проникающим лучам достигают 100 000 раз. Поражаемость ядра клетки под действием ионизирующих излучений, так же как и при действии других агентов, обусловлена высокой ранимостью хромосом. Локальное облучение хромосомы 60 протонами уже вызывает ее поражение. Высокая чувствительность ядра клетки к ионизации и другим воздействиям объясняется тем, что ДНК хромосом является самым чувствительным компонентом клетки по сравнению с белком и РНК цитоплазмы.
На различных стадиях митотического цикла чувствительность хромосом к облучению различна. Многие авторы считают, что наиболее высокая поражаемость хромосом наблюдается в интерфазе, так как при этом блокируется репликация молекулярного остова хромосом — ДНК.
Физиологическое состояние клеток обусловливает силу их реакции на облучение. Так, например, незрелые половые клетки более чувствительны, чем зрелые. Сухие семена растений более чувствительны, чем намоченные и т. д. Клетки разных тканей и клетки разного возраста одних и тех же тканей имеют различную чувствительность.
Чувствительности и устойчивость к радиации обусловлена наследственно. Глубина поражения облученных клеток зависит от генотипа организма клетки млекопитающих (и человека) менее устойчивы, чем клетки насекомых и растений. Одни виды животных и растений более чувствительны, чем другие. Полиплоидные клетки более устойчивы, чем диплоидные и гаплоидные того же вида. Путем селекции внутри вида можно создавать генотипы более и менее устойчивые к облучению.
Генетические причины, определяющие радиорезистентность, еще не установлены. Предполагается, что она обусловлена тем, что каждый генотип определяет различное соотношение химических ингредиентов в клетке, количество воды, степень и характер спирализации хромосом, а также продолжительность генерационного цикла (особенно S-фазы).
После облучения в ядрах клеток наблюдаются самые разнообразные обратимые и необратимые изменения: возникают клетки с гигантскими ядрами, многоядерные клетки, нарушается полярность при делении ядра, тормозится митотическая активность, происходит слипание хромосом (пикноз ядра) или их фрагментация и др. Во время деления облученной клетки в анафазе часто образуются хромосомные мостики,. что может быть следствием хромосомных перестроек или слипания хромосом. Нарушение нормального хода митоза под влиянием облучения может приводить к возникновению полиплоидных и гаплоидных клеток.
Поражение клеток организма может быть вызвано непосредственным облучением или косвенным воздействием через облученную среду, в которой помещены необлученные клетки, или дистантным действием ионизирующей радиации. Под дистантным действием имеют в виду возникновение патологических изменений ядерных структур в необлученной части ткани или органа под влиянием продуктов распада клеток облученной части того же организма. Продукты распада переносятся гуморальным путем.
Все три типа воздействия с разной силой вызывают наследственные изменения.
Генетический эффект облучения
Поскольку хромосомы обладают свойством репродуцироваться даже в измененном состоянии, различные изменения хромосом могут сохраняться в ряду клеточных делений и наследоваться.
Поэтому под генетическим эффектом ионизирующих излучений следует понимать любые изменения хромосом, связанные с возникновением мутаций. Обычно в большем количестве возникают летальные, семилетальные (понижающие жизнеспособность) и другие мутации, вызывающие гибель зигот. Такие мутации могут быть доминантными и рецессивными.
Если в сперматозоиде возникла мутация, то после слияния его с яйцеклеткой она перейдет в зиготу. Предположим, что в 10 из 1000 облученных сперматозоидов появились летальные мутации. После оплодотворения яйцеклеток из 1000 особей F1 около 1% будут нести данную мутацию в гетерозиготном состоянии. Если бы все эти летальные мутации оказались доминантными, то все 10 гетерозигот F1 погибли бы на определенных стадиях развития. Сравнивая количество отмирающих эмбрионов в потомстве облученных родителей с таковым в потомстве необлученных, можно установить частоту возникновения доминантных леталей, вызванных облучением половых клеток у родителей.
Рецессивные летальные мутации можно обнаружить лишь в последующих поколениях F2 и F3, т. е. после того, как они перейдут в гомозиготное состояние. Такой учет летальных мутаций прост и позволяет производить объективную количественную оценку частоты их возникновения.
На основании количественного учета мутаций была установлена зависимость частоты их возникновения от дозы облучения. Так, например, если облучить половые клетки дрозофилы рентгеновыми лучами в дозе 500 р, то соответственно этой дозе обнаруживается определенный процент рецессивных летальных мутаций, например около 0,5%. При повышении дозы облучения в 2 раза, т. е. до 1000 р, частота мутаций увеличивается также в 2 раза — до 1%. Облучение дозой в 3000 р повысит частоту мутаций в 6 раз, и т. д.
Многочисленные опыты с дрозофилой, кукурузой, ячменем и другими объектами позволили сделать вывод, что число точковых мутаций (n) возрастает прямо пропорционально дозе ионизирующего излучения (D): n = aND, где N — число исследованных хромосом и а — вероятность мутирования гена под влиянием дозы 1 р.
Указанную зависимость можно изобразить графически. На оси абсцисс откладывают дозы облучения, а на ординате — частоты обнаруженных мутаций. В этом случае график зависимости частоты мутаций от дозы облучения представит прямую линию (отсюда термин «линейная зависимость»).
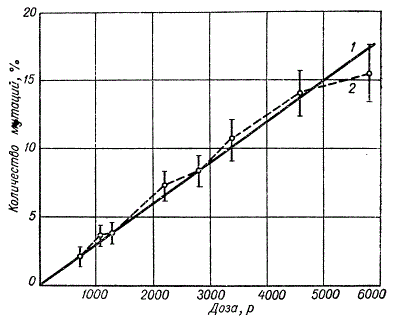
Зависимость частоты сцепленных с полом летальных мутаций от дозы ионизации
Продолженная влево на точку, соответствующую минимальной дозе, изученной в опыте, эта линия пройдет через нулевую точку координат. Это говорит ртом, что любая сколь угодно малая доза ионизирующей радиации будет приводить к повышению частоты мутаций. Вывод об отсутствии порога генетического эффекта ионизирующей радиации, сделанный на основе такой экстраполяции, получил теперь прямое подтверждение в опытах с малыми дозами. Существуют некоторые данные, позволяющие утверждать, что при любой малой дозе облучения повышение ее в 2 раза удваивает частоту возникновения мутаций.
Линейная зависимость частоты возникновения мутаций от дозы облучения навела на мысль о том, что мутация гена является следствием мономолекулярного акта, т. е. отдельной ионизации. Поэтому дальнейшие исследования сосредоточились в направлении изучения роли различных физических сторон действия ионизации на мутационный процесс, а именно влияния мощности (интенсивности) излучения — величины дозы в единицу времени, влияния плотности ионизации и возможности специфичного действия различных ионизирующих излучений.
Интересно отметить, что равные дозы ионизирующих излучений, различающиеся по интенсивности в 10 000 и даже 100 000 раз, вызывают мутации с одинаковой частотой.
Многочисленными работами Н. В. Тимофеева-Ресовского и его сотрудников в 30-х годах, а также ряда других исследователей было показано, что фракционирование одной и той же дозы не влияет на частоту возникновения мутаций. Так, облучение одной и той же дозой рентгеновых лучей (например, 1000 р), примененной непрерывно — в один сеанс облучения или фракционно — с интервалами в несколько часов, суток и даже недель, дает одинаковый процент мутаций.
Недавно Я. Л. Глембоцкий исследовал фракционированное и непрерывное действие малых доз рентгеновых лучей. Им было показано, что однократное облучение 20 р и та же доза, поданная в 4 сеанса (5 р Х 4) с перерывами в 1,5 ч, вызывают одинаковую частоту мутаций: соответственно 0,237 ± 0,071 и 0,218 ± 0,072. Приведенные опыты позволили сделать очень важный вывод о том, что генетический эффект ионизирующих излучений не зависит от скорости подачи дозы.
Далее было установлено, что частота точковых мутаций не зависит от жесткости рентгеновых лучей.
На основании этих и ряда других данных Н. В. Тимофеев-Ресовский в сотрудничестве с физиками еще в 1936 г. предложил гипотезу, вошедшую в литературу под названием «теории мишени». Эта гипотеза построена на признании принципа случайного попадания кванта энергии в ген. При попадании в ген кванта энергии, вызывающего ионизацию атома, ген мутирует с вероятностью, близкой к 1. Исходя из такого, чисто физического представления, он объяснял линейную зависимость частоты возникновения мутаций от дозы излучения. При этом предполагали, что ген мутирует необратимо по принципу «все или ничего». Ген принимался за мишень с определенным «чувствительным объемом». Поглощенная в пределах этого объема энергия «попавшего» кванта может «мигрировать» и вызвать изменение гена или разрыв хромосомы, который может привести к хромосомным перестройкам. Эта чисто физическая гипотеза, несомненно, сыграла положительную роль в изучении проблемы. Лежащие в ее основе положения и до сих пор успешно применяются в радиобиологии и радиационной генетике и известны теперь в литературе как «принцип попадания». Однако со временем «теория мишени» в той части, которая касалась механизма возникновения мутаций, вступила в противоречие с вновь открытыми фактами. Зависимость частоты возникновения мутаций от дозы облучения представляется теперь значительно более сложной, чем это казалось на первых порах.
Для более детального анализа влияния ионизирующих излучений на клетку и хромосомы необходимо познакомиться с их цитогенетическим эффектом. Данный эффект состоит в том, что под влиянием облучения возникают разнообразные структурные изменения хромосом, которые значительно отличаются от генных мутаций.
При действии на хромосомы ионизирующих излучений возникает большое количество хромосомных перестроек. Оказалось, что летальные мутации в большинстве своем связаны с хромосомными перестройками, которые можно обнаруживать генетическими и цитологическими методами. Так, В. В. Сахаров, используя метод анализа хромосомных перестроек на гигантских хромосомах слюнных желез дрозофилы, показал, что из 27 летальных мутаций, возникших под действием ионизирующего облучения, 21 оказалась хромосомной перестройкой, в то время как среди 71 мутации, возникшей спонтанно, была обнаружена лишь одна хромосомная перестройка, остальные 70 были точковыми генными изменениями.
Н. П. Дубинин с соавторами на огромном материале убедительно показали, что с увеличением дозы облучения (от 500 до 6000 р) увеличивается частота летальных мутаций, связанных с хромосомными перестройками.
Хромосомные перестройки возникают в результате двух актов: разрыва хромосомы и соединения разорванных концов. Разрыв образуется в результате непосредственного или опосредованного воздействия агента. Предполагается, что причиной разрыва хромосомы является ионизация атомов в молекулах ДНК. Соединение разорванных концов зависит от специфичности действия ионизирующего агента, от последующих изменений в клетке, возникших в результате его действия или влияния дополнительных факторов среды. Индуцированные разрывы в хромосомах могут происходить как одной точке, так и в двух, трех и т. д. Структурные изменения хромосомы, возникающие вследствие одного разрыва, называют одноударными хромосомными перестройками, структурные изменения вследствие двух одновременных разрывов — двухударными и т. д.
Одновременные перестройки происходят как в пределах одной хромосомы, так и в разных хромосомах. Известно, что каждая хромосома в зависимости от того, в какой фазе митотического цикла она находится, может состоять из различного числа хромосомных нитей: одной — от анафазы до ранней интерфазы, т. е. до редупликации, и двух-четырех — после редупликации. Поэтому при действии ионизирующих излучений на хромосомы в разных фазах митотического цикла разрывы происходят либо в одиночной хромосомной нити (хромосомные разрывы), либо в двух (хроматидные), либо в четырех (пол у хроматидные). Эти разрывы могут быть в тождественных (симметричных) и нетождественных (асимметричных) участках хроматид и полухроматид. Так как разорванные концы обладают свойством соединяться, то произошедшие одиночные и двойные разрывы в одиночной или двойной хромосомной нити обусловливают различные хромосомные перестройки. Последние выявляются на цитологических препаратах в метафазе и анафазе в виде различных конфигураций.
Вследствие различных комбинаций при перестройке хромосом возникают концевые, внутренние и кольцевые дицентрические делеции и взаимные транслокации. При хроматидных разрывах в зависимости от их числа (одинарные или двойные разрывы) возникают хроматидные и изохроматидные делеции, внутри- и межхромосомные обмены, которые могут быть симметричными и асимметричными.
Необходимо подчеркнуть, что действие ионизирующих излучений на возникновение хромосомных перестроек сначала было обнаружено генетическими методами. Впоследствии этот эффект был подтвержден цитологическими методами на хромосомах соматических клеток корешков растений, эмбриональных тканей и костного мозга животных и человека, а также, на гигантских хромосомах слюнных желез дрозофилы.
Хромосомные и хроматидные перестройки, возникающие на разных стадиях митоза, нарушают нормальный его ход и могут приводить к гибели клетки. Однако происшедшие разрывы могут частично воссоединиться таким образом, что восстановится исходная структура. Вследствие этого истинная частота возникающих хромосомных перестроек и частота аберраций, обнаруживаемых генетическими и цитологическими методами, могут далеко не совпадать. Поэтому учитываемая частота возникновения хромосомных перестроек в зависимости от дозы облучения и условий облучения отражает истинную частоту их возникновения лишь относительно.
Частота возникновения разных типов хромосомных перестроек находится в различной зависимости от дозы ионизирующей радиации. Частота хромосомных перестроек, происходящих в результате одиночного разрыва, находится в линейной зависимости or дозы. Частота же двух независимых одновременных разрывов, вызванных ионизацией и, соответственно, основанных на них двухударных перестроек, возрастает пропорционально квадрату дозы вследствие того, что вероятность одновременного возникновения двух независимых событий равна произведению вероятностей.
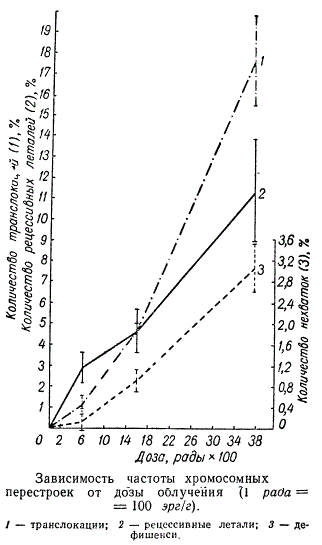
Такая зависимость возможна при условии, что разные дозы ионизации вызывают с одинаковой вероятностью разрывы и воссоединение разорванных концов в расчете на единицу дозы.
Зависимость между возникновением межхромосомных перестроек и дозой облучения не является линейной. Однако характер ее еще недостаточно изучен. Трудность количественного анализа возникновения различных хромосомных перестроек в зависимости от дозы заключается в том, что во время воздействия могут возникать истинные и потенциальные разрывы хромосом. Последние в зависимости от условий, складывающихся в клетке после облучения, могут реализоваться в истинные разрывы или совсем не реализоваться. Открытые концы в истинных разрывах, возникших в момент облучения, воссоединяются в новой комбинации или в исходное состояние в течение какого-то времени после взрыва.
На основании различных опытов складывается представление, что способность разорванных концов хромосом соединению в новой комбинации или воссоединению исходной структуры зависит от фазы митотического и мейотического циклов клетки, специфики объекта, характера излучения и от биохимических условий микросреды.
Так, прямыми цитологическими исследованиями — подсчетом клеток с нарушенными хромосомами — показано, что возникновение хромосомных перестроек зависит от плотности ионизации, облучения с меньшей энергией и большей плотностью ионизации не эффективны в вызывании хромосомных перестроек. Нейтроны, обладающие, например, энергией 7,5 Мэв, оказываются более эффективными в вызывании хромосомных аберраций, чем нейтроны с энергией 15 Мэв.
В опытах с рентгеновыми лучами показано, что их эффективность в вызывании хроматидных перестроек зависит от жесткости лучей: наиболее эффективные лучи с длиной волны 4,1 Å, менее эффективны — 0,15 Å. Еще менее эффективны γ-лучи. В общей форме можно сказать, что корпускулярные излучения — быстрые нейтроны и α-частицы — вызывают хромосомные перестройки чаще, чем электромагнитные излучения. Это различие объясняется разницей в плотности ионизации, которую они производят.
Вследствие того, что хромосомные перестройки возникают в два акта и при этом реализуется лишь незначительная часть истинных разрывов, а остальные либо восстанавливаются, либо элиминируются, обнаруживаемая частота структурных изменений хромосом оказывается зависимой не только от дозы, но и от многих других факторов. На частоту возникновения обнаруживаемых перестроек влияет и фракционирование облучения.
Под влиянием ионизирующего облучения клеток увеличивается также возможность нерасхождения хромосом, в результате чего появляются гетероплоидные ядра (анеуплоидные). Неравномерное распределение хромосом в митозе в облученных клетках, по-видимому, может быть следствием двух причин: первичной — из-за разрушения ахроматинового веретена, растаскивающего хромосомы к полюсам деления, либо вторичной — из-за слипания хромосом, вызванного произошедшими хромосомными перестройками. Возникновение нерасхождения хромосом зависит от дозы ионизации, стадии развития половых клеток, возраста организма и его генотипа.
Действие облучения ведет также к значительному усилению кроссинговера.
Для изучения роли прямого и непрямого действия мутагенов необходимо исследование химических процессов, разыгрывающихся в клетках в результате ионизации. Уже в начале нашего столетия было показано, что облучение воды ведет к ее разложению (радиолиз). Позднее было установлено, что в результате радиолиза воды образуется перекись водорода. При этом выход ее зависит от вида излучения и от плотности ионизации.
Наличие в живых тканях большого количества воды и тот факт, что ионизация вызывает разложение воды, навело исследователей на мысль о роли кислорода в мутационном процессе, индуцированном ионизирующей радиацией. Рядом исследователей было показано, что облучение дрозофилы в атмосфере чистого кислорода повышает частоту мутаций, а облучение в атмосфере азота снижает частоту обнаруживаемых мутаций. Бескислородная атмосфера оказывается в некоторой мере защитной при ионизирующем облучении клетки. Явление увеличения частоты мутаций при действии ионизирующей радиации в присутствии кислорода было названо кислородным эффектом. Влияние кислородного эффекта на возникновение хромосомных перестроек подтвердилось в опытах на растениях и животных. Кислород увеличивает чувствительность хромосом к облучению. Причем повышение его концентрации во время облучения от 0 до 21% линейно увеличивает число хромосомных перестроек; дальнейшее повышение концентрации кислорода оказывается менее эффективным.
Бескислородная среда после облучения ослабляет эффект ионизации. Наличие кислорода в среде после облучения, напротив, усиливает этот эффект. Эти явления подтверждают положение
о том, что хромосомные перестройки возникают в результате обратимого нарушения в ядре клетки, вызванного облучением. Подобные факты все более склоняют исследователей к тому, что мутации являются следствием вторичных процессов в хромосомах типа депатурации ДНК и иуклеопротеидов, вызываемых ионизирующими излучениями.
Итак, все виды ионизирующих излучений вызывают глубокие изменения хромосом, которые благодаря репродукции воспроизводятся в митозе и могут сохраняться в ряду последующих клеточных делений.
Влияние ультрафиолетовых лучей
Ультрафиолетовые лучи (УФ) способны возбуждать электронные оболочки, в результате чего образуются фотоэлектроны, которые вызывают различные химические реакции, приводящие к мутациям.
Благодаря большой длине волны УФ-лучи легко абсорбируются поверхностными тканями организма и плохо проникают во внутренние ткани. УФ-лучи более эффективны в вызывании мутаций у одноклеточных организмов, так как легко достигают ядра клетки.
В настоящее время мутагенное действие УФ-лучей доказано для всех организмов, у которых возможно непосредственное облучение генеративной ткани или половых клеток. Они вызывают все виды мутаций. Частота мутаций зависит от дозы облучения, но до определенного предела, выше которого частота мутаций не повышается, а у ряда организмов даже снижается.
Но самой интересной особенностью действия УФ-лучей является то, что наибольшая эффективность их связана с определенным спектром длин волн, а именно с областью от 2500 до 2800 Å.
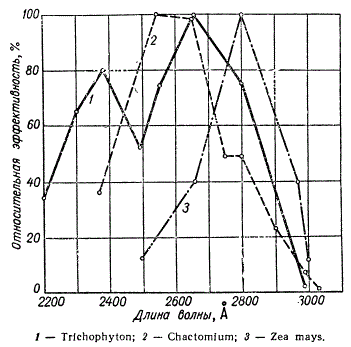
Эффективность вызывания мутаций УФ-лучами с различной длиной волны у различных организмов
Это открытие явилось важным моментом в понимании природы наследственной изменчивости. Дело в том, что нуклеиновые кислоты, составляющие основу хромосом, поглощают волны как раз этой части спектра, т. е. около 2600 Å. Лучи с такой длиной волны наиболее эффективно разрушают ДНК, и, по-видимому, этим объясняется высокая чувствительность к ним ядра клетки. Волны этой части спектра УФ, вероятно, именно поэтому обладают наибольшим бактерицидным действием.
Механизм действия УФ на возникновение мутаций представляет большой интерес. Исследование действия УФ на мутационный процесс у бактерий показал, что облучение даже не самого объекта, а лишь среды, на которой развиваются клетки, может индуцировать мутации. Так, у кишечной палочки Escherichia coli мутации, повышавшие устойчивость к стрептомицину и пенициллину, возникали в 10—500 раз чаще, чем спонтанные, если бактерии развивались на облученной УФ среде. Оказывается, что если предварительно облучить УФ среду, содержащую аминокислоты, а затем поместить на нее конидии нейроспоры, то частота возникающих мутаций у них также увеличивается. Облученная среда в данном случае действует как мутаген даже спустя несколько часов после облучения.
Облучение среды УФ вызывает в ней образование активных соединений типа перекиси водорода и органических перекисей, которые оказываются мутагенами. Это доказывается следующим образом. Если предварительно в облучаемой среде, в которую затем помещаются микроорганизмы, или в самих клетках понизить содержание кислорода, а, следовательно, и уменьшить образование перекиси водорода, то частота мутаций не увеличивается. Того же эффекта можно добиться путем прибавки фермента каталазы, разрушающего перекись водорода. Следовательно, перекись водорода оказывается мутагеном, а каталаза — антимутагеном.
Часть спектра УФ-лучей обладает свойством фотореактивации. Это явление заключается в том, что лучи в пределах волн длиной 3650—4900 Å (видимая часть спектра — синий свет) подавляет мутагенный эффект активной части спектра УФ. И более того, они оказывают подавляющее влияние на действие ионизирующих излучений, так как усиливают активность ферментов, в частности цитохромоксидазы, понижающих концентрацию различных перекисей. Указанная часть спектра (3650—4900 Å) соответствует спектру поглощения цитохромоксидазы. В общем, надо сказать, что УФ-лучи являются мощным мутагеном.