Водный режим почв рассматривается как один из важнейших показателей их лесорастительных свойств.
Древесные породы характеризуются разной требовательностью к условиям водного питания и поглощают из почвы неодинаковое количество влаги. Показателем расхода влаги деревьями является величина транспирации — физиологического испарения воды листьями (хвоей) растений. Высокопродуктивные насаждения расходуют на транспирацию больше влаги.
Величина транспирации зависит от запасов влаги в почве. Наибольший расход влаги на транспирацию соответствует некоторому среднему запасу влаги в почве (например, для сосняка — около 180 мм за вегетационный период в слое почвы 100 см). При запасах выше и ниже этого значения расход влаги древостоем меньше.
Величина транспирации изменяется в зависимости от возраста древостоя. Д ля сосновых насаждений расход влаги на транспирацию максимален в возрасте 20—40 лет. Установлена положительная зависимость между величиной годичного текущего прироста древостоя и расходом влаги на транспирацию.
В качестве интегральной экологической характеристики отношения древесных пород к почвенной влаге выдающийся русский лесовод Г. Ф. Морозов предложил использовать понятия «потребность» и «требовательность». Потребность — это количество влаги, которое необходимо для жизнедеятельности растений (нормального хода фотосинтеза, дыхания, обмена веществом между органами и т. д.). Требовательность — это отношение растений к условиям водного режима почв, их способность взять необходимое количество влаги в тех или иных почвенных условиях.
Запасы продуктивной влаги, необходимые для роста и развития растений, неодинковы в разных типах почв.
По Г. Ф. Морозову (1926), потребность древесных пород во влаге может быть охарактеризована как совокупность признаков: интенсивности, продуктивности и экономности транспирации. Первая величина измеряется количеством воды на единицу площади листьев, вторая — на количество всего органического вещества прироста дерева, третья — на количество начального запаса влаги во всем растении.
Так, потребность во влаге сосны, ели, пихты относительно невелика. Но эти породы сильно различаются по требовательности к содержанию влаги в почве. Сосна (ксерофит с развитой глубинной составляющей корневой системы) нетребовательна к почвенной влаге, тогда как ель и пихта (мезофиты с преимущественно поверхностной корневой системой) более требовательны к содержанию влаги в почве. Сосна и ель близки по интенсивности испарения, однако сосна в два раза экономнее расходует влагу на транспирацию. Ели необходимо в течение суток дважды восполнять запас влаги в своих органах. Следовательно, запасы продуктивной влаги в почвах для ели должны быть выше, чем для сосны.
С особенностью водного режима почв связан тип местообитания леса, характеризуемый количеством продуктивной влаги. Водный режим почв определяется соотношением приходной и расходной частей водного баланса и водоудерживающей способности почв.
Сухие и крайне сухие местообитания
Условия этого типа местообитаний характерны для песчаных малогумусных грубоскелетных почв на глубоких песках с очень низкой влагоемкостью и водовместимостью.
Содержание влаги в почвах сухих местообитаний находится в диапазоне от ВРК до ВЗ. Принято считать, что влажность завядания древесных пород соответствует значению двойной максимальной гигроскопичности. Среди древесных пород в таких местообитаниях произрастают сосны и дубы низкого бонитета, можжевельник, грабинник, а также степные кустарники. В напочвенном покрове преобладают лишайники, бессмертник и другие ксерофиты. При сплошных рубках наблюдается формирование степного растительного покрова.
При наличии в почвах прослоек (псевдофибр), а также утяжелении гранулометрического состава за счет пылеватых фракций, увеличивающих водоудерживающую способность почвы, достигается II класс бонитета сосны и III—IV — дуба. К ним могут примешиваться мезофиты — липа, ясень, бук III—IV классов бонитета.
Различные древесные породы обладают неодинаковой способностью извлекать влагу из почвы. Почвенная засуха является результатом атмосферной и сопровождающей ее сильной транспирации растений. Корни деревьев, поглощая влагу, исчерпывают физиологически доступные ей запасы. Прочно связанная, недоступная растениям влага составляет в песках 0,2—0,5%, в суглинках — 5—10%. Засухе подвержены большие площади земель. Научные основы борьбы с засухой были заложены в трудах В. В. Докучаева, Г. Н. Высоцкого и Г. Ф. Морозова.
Один из эффективных способов повышения плодородия почв, борьбы с опустыниванием и деградацией земель — агролесомелиорация. Основу агролесомелиорации составляют полезащитные лесные насаждения и лесомелиоративные комплексыв в степных и полупустынных ландшафтах. За всю историю защитного лесоразведения на территории России засеяно 5,2 млн га. Главные задачи агролесомелиорации — улучшение водного режима, обустройство земель и защита почв от водной и ветровой эрозии. Для полезащитных лесов засушливого юга главным является не столько прирост древесины, сколько сохранение их жизнеспособности. Для создания полезащитных лесных полос используются устойчивые к засухе породы деревьев, такие как сосна, дуб, мелколиственный берест, белая акация, американский клен, серебристый лох.
Свежие местообитания
Такие местообитания встречаются в разных типах водного режима. При КУ = 1 и непромывном типе водного режима свежие местообитания характерны для серых почв и буроземов. На песчаных почвах свежие местообитания формируются при залегании грунтовых вод на глубине 2—4 м. На суглинистых почвах они обеспечиваются за счет высокой влагоемкости при уровне грунтовых вод глубже 5 м. Наименьшая влагоемкость суглинистых почв в 4 — 5 раз больше, чем песчаных. В сухом климате при КУ < 1 свежие местообитания возникают в почвах понижений, а в средних и нижних частях склонов — за счет дополнительного притока влаги.
Во влажном климате при КУ > 1 свежие местообитания характерны для хорошо дренированных почв. В таких местообитаниях содержание продуктивной влаги оптимально для достижения высшей производительности многих древесных пород: сосны, березы бородавчатой, граба, клена, ясеня, бука, пихты, лиственницы. В свежих местообитаниях в напочвенном покрове сосновых лесов господствуют зеленые мхи, брусника, грушанка, орляк. В напочвенном покрове дубрав преобладают ясменник, сныть и другие характерные растения.
При изменении влажности от НВ до ВУЗ происходит быстрое снижение транспирации и сокращение прироста древостоя. При уменьшении влажности до ВЗ и ниже расход влаги на транспирацию сокращается до чрезвычайно низких величин вследствие прочного удержания почвой остаточной влаги.
Влажные местообитания
Они формируются в песчаных почвах при глубине грунтовых вод от 1 до 2 м, а в суглинистых почвах — от 3 до 5 м. Корнеобитаемый слой в этих почвах находится в зоне капиллярной каймы. При более глубоком залегании грунтовых вод влажные местообитания образуются при двучленном строении почвообразующих пород, когда песчаный верхний слой подстилается суглинистой породой на глубине 80—140 см. В таких почвах образуется верховодка, которая служит источником дополнительной влаги для растений.
Влажные местообитания оптимальны для ели, черешчатого дуба, пушистой березы, ивы, тополей. В еловых лесах в напочвенном покрове преобладают зеленые мхи, кукушкин лен, черника, кислица. Под широколиственными лесами в напочвенном покрове распространены папоротник, сныть, медуница. Встречаются как мезофиты, так и мезогигрофиты и даже гигрофиты.
Сырые местообитания
Данный тип местообитаний характеризуется избыточным увлажнением для большинства пород деревьев. Они образуются, когда зеркало грунтовых вод в песчаных почвах находится на глубине 0,5—1,0 м, а в суглинистых — 1—3 м. На протяжении большей части вегетационного периода со-держание влаги в почвенном профиле превышает НВ. Сырые местообитания характерны для суглинистых почв территории лесной зоны с КУ > 1 вследствие низкой водопроницаемости текстурного горизонта и дополнительного поступления влаги с боковым внутрипочвенным стоком.
В насаждениях сырых местообитаний господствуют ель, сосна, дуб, осина, береза. Будучи мезофитами, эти деревья в условиях избытка влаги в профиле почв развивают приповерхностные корневые системы, что ведет к потере ветроустойчивости, образованию характерного для влажных лесов микрорельефа из-за корневых выворотов.
Сырые местообитания оптимальны для мезогигрофитов — серой ольхи, черемухи, серой ивы. В сырых местообитаниях вырубки заболачиваются и переходят в категорию болот за относительно короткое время.
Влажные местообитания лесных болот
Для них свойственны постоянно избыточные застойные или слабопроточные режимы увлажнения торфяных болотных почв. Грунтовые воды выходят на поверхность почвы весной и сохраняют высокий уровень в вегетационный период. На сфагновых болотах уровень грунтовых вод обычно не опускается ниже 30—50 см, в ольшаниках — до 50—80 см.
Постоянное избыточное увлажнение отрицательно влияет на рост большинства древесных пород. Только черная ольха представляет собой настоящий гигрофит. Наилучшие условия требовательная к минеральному питанию ольха находит в торфяных эутрофных почвах.
Среди древесных пород на торфяных олиготрофных почвах преобладают низкоствольные сосна и береза ниже V класса бонитета. Эти породы приурочены к кочкам и развивают мелкую горизонтальную корневую систему в слое 20—30 см.
Пойменный тип местообитания
Аллювиальные почвы, характеризующиеся контрастным режимом увлажнения, образуют пойменный тип местообитания. На протяжении одного вегетационного периода пойменные территории в зависимости от водного режима реки могут переходить из состояния полного затопления в паводковый период в состояние дефицита влаги на песчаных прирусловых валах в меженный период.
Разные породы деревьев в неодинаковой степени выносят затопление. Древовидная ива (особенно черная и серебристая), тополь способны сохранять жизнеспособность при уровне полых вод до 5—6 м выше шейки корня в течение 2—3 мес. Ольха и вяз выдерживают затопления до 1—1,5 месяцев, дуб — до 1 мес, береза, ель, сосна — до 2—3 недель. Полностью не выносят затоплений бук, граб, пихта.
Влияние увлажнения и аэрации почв на производительность древесных насаждений
Это влияние показано на примере экологического профиля сосняков через боровой комплекс на озовой возвышенности. Почвы на профиле сменяются последовательно от подбуров в сухом и свежем борах (А1, А2) через подзолы и подзолы глеевые во влажном и сыром (А3, А4) до торфяно-глеевых в сыром (А5).
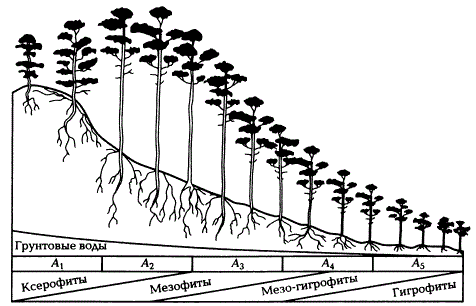
Боровой экологический ряд сосняков на склоне дюны от сухого бора А1 на ее вершине до сосны по болоту А5 у ее подножия (по Н.П. Ремезову, П.С. Погребняк, 1965)
Боровой комплекс на профиле бедных песчаных почв формируется под влиянием разных степеней их увлажнения. На верхней части бугра произрастает лишайниковый бор из чистой сосны III—IV классов бонитета (А1). Ниже по склону он переходит в сосняк зеленомошно-брусничный I—II класса бонитета (А2). Далее свежий бор сменяется влажным бором зеленомошно-черничным III класса бонитета (А3). В нижней части склона он сменяется сосняком долгомошным IV класса бонитета с черникой, голубикой, багульником, сфагнумом (А4). Экологический ряд заканчивается болотом, расположенным у подножия склона с редкой сосной V класса бонитета (А5).
Влажность почвы в данном случае является ведущим экологическим фактором производительности древостоя. На отрезке профиля от сухого бора до влажного бонитет сосняков увеличивается. Это связано с дополнительным поступлением влаги к корнеобитаемому слою за счет капиллярного подтока из грунтовых вод, более насыщенных минеральными веществами.
Главным фактором высокой продуктивности древостоев является устойчивое обеспечение их продуктивной влагой в течение всего их жизненного цикла.
Ниже по склону водообеспеченность возрастает. Однако в самой нижней части склона, где уровень грунтовых вод достигает корнеобитаемой зоны, резко ухудшается аэрация почвы. При избыточном увлажнении в почве возникают восстановительные условия и дефицит кислорода, появляются токсичные соединения. Это приводит к существенным изменениям в лесорастительном эффекте увлажнения. Бонитет сосняков в местообитаниях влажного сырого бора и болота сильно падает.
Как постоянный избыток, так и постоянный недостаток влаги оказывают сходное влияние на рост деревьев. В обоих случаях корневая система «уплощается», становится приповерхностной, что приводит к уменьшению площади питания растений и, следовательно, ухудшению его пищевого режима. Этому способствуют и косвенные факторы: снижение биологической активности почв, уменьшение количества опада.
Устойчивость водного питания растений тесно связана с уровнем грунтовых вод и водоподъемной способностью почв. При отсутствии связи водного режима почв с грунтовыми водами тип местообитания зависит от объема и характера поступления осадков, их перераспределения в почвенном профиле, водоудерживающей способности почв и степени дренированности территории. Различные типы местообитаний, обусловленные водным режимом почв, могут соседствовать друг с другом, обнаруживая тесную связь со структурой почвенного покрова. При сложном строении почвенного покрова в лесной зоне наблюдается мозаичная структура местообитаний древесных пород.
На развитие корневых систем древесных растений влажных местообитаний большое влияние оказывает аэрация почв. В зону капиллярного подъема грунтовых вод, где содержание воздуха в почве меньше 30 %, корни лиственных пород и ели заходят лишь на небольшую глубину. Она ограничена пределами сезонного колебания верхней границы капиллярной каймы. Корни сосны не выносят такого избытка влаги и ежегодно отмирают.
Влияние аэрации на корни деревьев обусловлено общим количеством воздуха в почве, содержанием в ней кислорода, углекислого газа, сероводорода, метана. Большинство растений бывает угнетено при содержании в почвенном воздухе менее 9—12% кислорода и более 1 % углекислого газа.
Между типом водного режима и морфологическими свойствами почв существует тесная связь. Поэтому морфологические признаки почвенного профиля могут быть индикатором местообитания по степени увлажнения. К ним в лесной зоне относятся мощность органогенного горизонта, строение лесной подстилки, мощность подзолистого горизонта, наличие и формы новообразований, признаки оглеения (сизая окраска и положение глеевого горизонта в профиле), глубина уровня грунтовых вод или верховодки, а также Запах сероводорода в почвенном разрезе.
В степной зоне индикаторами местообитаний могут быть признаки оглеения в профиле почв, граница капиллярной каймы, уровень грунтовых вод, глубина иллювиального карбонатного горизонта, форма новообразований карбонатов.
Морфологическая диагностика потенциального плодородия лесных почв
Показателями относительно высокого потенциального плодородия почв являются полноразвитый почвенный профиль при слабоуплотненном сложении. Мощность гумусового горизонта более 10 см, муллевый тип гумуса, отсутствие признаков постоянного переувлажнения (доля сизых пятен до 30 % от площади профиля), положение уровня грунтовых вод относительно поверхности земли в вегетационный период глубже 2 м в суглинистых почвах, мощность лесной подстилки до 10 см, двучленное строение почвенного профиля (подстилание песка супесью или суглинком на глубине до 120 см), наличие тонких (до 1 см) уплотненных супесчаных или ожелезненных прослоек в профиле песчаных почв.