С помощью хромосомных перестроек возможно:
- изучать взаимодействие генов при изменении их положения в хромосоме;
- выяснять влияние расположения эухроматинового и гетерохроматинового материала на фенотипический эффект гена;
- исследовать межхромосомные отношения в генотипе организма;
- получать новые группы сцепления.
Иначе говоря, ту структуру кариотипа и генотипа видов, которая отрабатывалась в ходе эволюции в течение сотен тысяч и миллионов лет, генетик имеет возможность перестроить в течение нескольких поколений. С помощью хромосомных перестроек можно создавать новые системы генотипов.
Хромосомные перестройки, происходящие как внутри одной хромосомы, так и между негомологичными хромосомами, являются очень важным механизмом перекомбинации генов внутри хромосомного набора каждого вида.
Из изложенного должно быть ясно, что перестройки хромосом могут изменять поведение хромосом в мейозе, действие генов, свойств доминирования генов, характер рекомбинации генов, гаметогенез и т. д. Поскольку естественный отбор контролирует все процессы в организме, очевидно, что потомство организмов с разными хромосомными перестройками будет иметь разные шансы на выживаемость.
Мы уже говорили о том, что явление гомологичной наследственной изменчивости объясняется в ряде случаев происхождением видов на основе хромосомных перестроек исходного кариотипа. Однако следует помнить, что когда формулировался закон гомологических рядов, то еще не было достаточного количества фактов о наличие хромосомных перестроек, и закон разрабатывался на основе фенотипической классификации наследственной изменчивости.
Теперь в генетике накоплено большое количество фактов, дающих основание признать, что одним из основных механизмов, обусловливающим возникновение гомологических рядов мутаций у близких видов, является процесс хромосомных перестроек. Транслокации, инверсии, дупликации и полиплоидия в процессе дифференциации вида на расы, подвиды и новые виды играют роль изолирующих факторов одной группы индивидуумов от другой. Указанные хромосомные перестройки вызывают нескрещиваемость особей в популяции, а также снижение плодовитости и жизнеспособности зигот вследствие нарушения генного баланса. Но в случаях возникновения жизнеспособной формы, гомозиготной по транслокации, инверсии или дупликации, она может оказаться приспособленной к определенным условиям существования и свободно размножится, а затем обособится в новый вид. У этого нового вида сохраняются прежние гены, но либо они окажутся в других группах сцепления, либо в иной последовательности расположения. Такие гены могут мутировать в том же направлении, что и у исходного вида, и таким образом обусловливать возникновение гомологичных рядов мутаций. Как показывают генетические исследования родственных видов, особенно в роде Drosophila, их генетические системы оказываются очень сходными, а различия касаются главным образом расположения отдельных генов.
Роль хромосомных перестроек важна и для эволюции генотипа. Как было показано, в результате транслокаций, дупликаций и инверсий гены вследствие эффекта положения изменяют характер Доминирования. Если полезная мутация гена является рецессивной, то с помощью эффекта положения она может проявиться в гетерозиготном состоянии и стабилизироваться в жизни вида. Значение транслокаций особенно велико в переносе отдельных участков аутосом на половые хромосомы. Эти перестройки являются важным фактором в определении нескрещиваемости видов животных.
Как мы уже знаем, Y-хромосома чаще состоит из гетерохроматина и генетически мало активна. Но у разных животных это проявляется в разной степени. Так, при общей малой активности Y-хромосомы дрозофилы один из ее участков, однако, является гомологичным участку в Х-хромосоме. В Y-хромосоме человека такой участок значительно длиннее, то же самое имеет место и у растения меландриум.
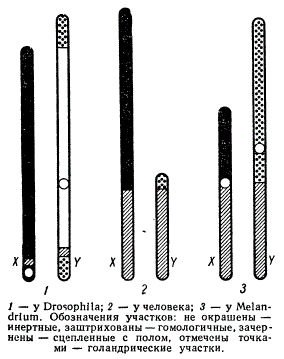
Сравнительное распределение генетически активных и инертных участков X- и Y- хромосом
В X- и Y-хромосомах могут быть как гомологичные участки, так и негомологичные, т. е. в Х-хромосоме всегда имеется свой, характерный для нее, район, отсутствующий в Y-хромосоме; гены в этом районе будут наследоваться сцепленно с полом. В Y-хромосоме также имеется район, не представленный в Х-хромосоме. Признаки, определяемые генами этого района (голандрическими генами), наследуются только по мужской линии при гетерогаметичности мужского пола.
Гомологичные и негомологичные участки в половых хромосомах, очевидно, могли произойти также посредством хромосомных перестроек. Об этом говорит тот факт, что у многих видов животных число гетероморфных хромосом сильно варьирует.
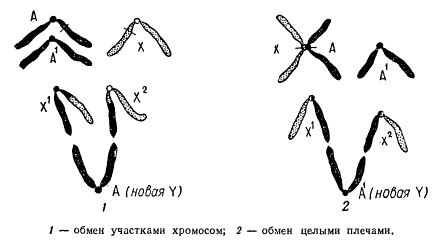
Схема образования гетероморфных хромосом у гетерогаметного пола (X0) в результате транслокации между X- хромосомой и аутосомой
Схематически показан ход возможного обмена фрагментами между Х-хромосомой и аутосомой у гетерогаметного пола Х0 в результате транслокации. Вследствие такой перестройки образуется новый тип гетерохромосом X1 X2 Y. Цитологический анализ мейоза у таких форм показывает образование тривалентов, что подтверждает правильность предположения о происхождении гетероморфных хромосом посредством транслокаций.
Цитологический анализ у некоторых родственных видов животных показывает, что гаплоидные числа хромосом в их наборе различаются, при этом, отдельные хромосомы могут нести гомологичные участки. У некоторых родственных видов дрозофилы известны 3 пары хромосом (D. willistoni), у других — 4 пары (D. melanogaster и D. americana), у третьей группы — 6 пар (D. virilis).
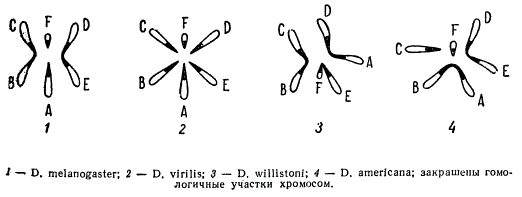
Гаплоидные наборы хромосом некоторых видов дрозофилы
Хромосомы могут превращаться в двуплечие хромосомы и, наоборот, может изменяться также и число хромосом в наборе.
Результаты исследования хромосомных перестроек убеждают во-первых, в наличии линейной дискретности хромосом и, Во-вторых, в том, что генотип представляет целостную систему, не сумму отдельных генов.
Рассмотрение хромосомных перестроек приводит к выводу, что они:
- лежат в основе изменений групп сцепления генов;
- изменяют характер наследования признаков и свойств в поколениях;
- изменяют проявление и взаимодействие генов;
- являются не только источником наследственной изменчивости комбинативного характера, но и механизмом преобразования генотипа и кариотипа в процессе эволюции;
- свидетельствуют о том, что многие гены, считавшиеся «классическими» точковыми мутациями, оказываются или дупликациями, или делециями, или инверсиями.
Хромосомные перестройки служат методом цитогенетической локализации генов в хромосомах, методом исследования механизма мейоза и тонкого картирования генов. Они могут быть использованы в практических целях для изменения групп сцепления генов, определяющих хозяйственно-ценные признаки.