Как ни прост общий план строения тела растения, все же оно расчленено на органы, имеющие ярко выраженную специализацию и развивающиеся в резко различных условиях среды.
При этом органы, выполняющие разные функции, например лист и корень, пространственно разобщены и удалены друг от друга иногда на значительное расстояние. Непрерывная и экономичная переброска строительных и энергетических материалов от одного органа к другому — необходимое условие нормальной жизнедеятельности растения.
Целостность растительного организма, тесная связь и взаимодействие органов находят свое выражение в развитии единой для всего растения проводящей системы. Основной единицей проводящей системы растения является ксилемно-флоэмный пучок, представляющий собой непрерывную транспортную магистраль, соединяющую органы различного функционального значения. Строение проводящего аппарата в отдельных звеньях меняется в зависимости от структурной и функциональной специализации органов. Так, для корня как органа поглощения минеральных веществ из почвы физиологически выгодным является радиальное расположение флоэмы и ксилемы в непосредственной близости обеих тканей к тканям коры. Для листа и стебля наиболее целесообразно расположение обеих тканей в коллатеральных или биколлатеральных пучках. Различное размещение проводящих тканей в корне и стебле выработалось в процессе эволюции почти у всех групп высших растений м особенно отчетливо выражено у двудольных.
Переход структуры корня в структуру стебля осуществляется в проростках между корнем и семядолями. В простейшем случае корень имеет два луча протоксилемы, располагающихся в плоскости, проходящей через середину семядолей. Флоэма в этом случае дифференцируется в межсемядольной плоскости, т. е. флоэмные и ксилемные группы располагаются крестообразно. Что же касается семядолей, то в них находятся типичные коллатеральные пучки с протоксилемой, дифференцирующейся на внутренней стороне прокамбиального тяжа. Различие в расположении тканей таким образом весьма существенное, и в то же время проводящие ткани непрерывно продолжаются из одного органа в другой. Детальное изучение процесса дифференциации тканей в проростках позволяет понять, каким образом осуществляется перестройка структуры проводящего аппарата при переходе от корня к стеблю. На рисунке изображена серия поперечных срезов через «зону перехода» у 5-дневных проростков томатов. Среди дифференцированных элементов ксилемы мы видим только одиночные сосуды, относящиеся к протоксилеме, а отчасти к метаксилеме.
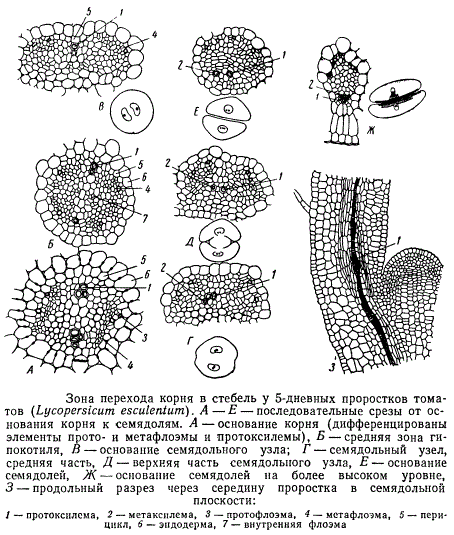
На протяжении гипокотиля и семядольного узла положение сосудов протоксилемы по отношению к перициклу изменяется: они постепенно все дальше отходят от перицикла, дифференцируясь во все более глубоких слоях прокамбия. В основании семядолей элементы протоксилемы занимают самое внутреннее (эндархное) положение в дифференцирующемся прокамбиальном тяже. Более последовательно изменение в положении элементов протоксилемы показано на рисунке, изображающем серию срезов семядольного узла.
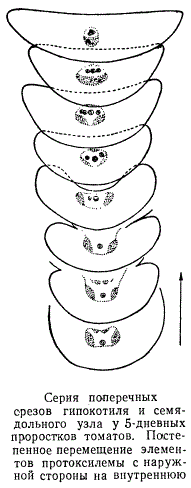
Как можно видеть, самый первый по времени созревания центральный сосуд постепенно углубляется, отходя от перицикла и смещаясь к внутренней стороне прокамбиального тяжа, пока окончательно не займет эндархного положения.
Процесс постепенного «углубления» элементов протоксилемы можно наблюдать и на продольных срезах, проведенных строго через медианную плоскость семядоли. Цепочка члеников сосуда, сохраняя непрерывность, постепенно смещается к внутренней стороне семядоли: каждый новый членик дифференцируется во все более глубоких слоях прокамбия.
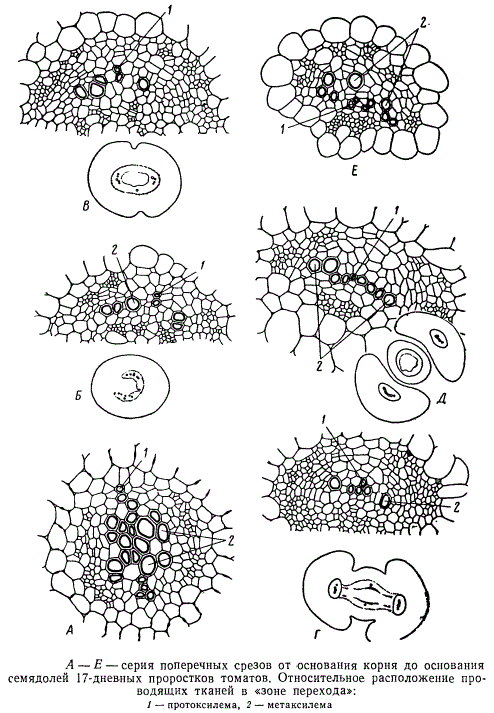
Локализация первых элементов протоксилемы определяет ход последующей дифференциации проводящих тканей в прокамбии. В корне элементы метаксилемы дифференцируются внутрь от сосудов протоксилемы, занимающих крайнее положение в центральном цилиндре, а в семядолях, наоборот, дифференциация элементов метаксилемы идет центробежно — кнаружи от протоксилемы. В гипокотиле расположение элементов метаксилемы в значительной мере определяется также развитием центральной паренхимы, отодвигающей к периферии весь комплекс проводящих тканей. Метаксилема дифференцируется в этой зоне большей частью по сторонам от сосудов протоксилемы, занимая вместе с тем более глубокое положение в центральном цилиндре. В семядольном узле сосуды метаксилемы постепенно «подтягиваются» к протоксилеме, оказываются на одном уровне с нею, а затем уже в основании семядоли элементы метаксилемы дифференцируются кнаружи от протоксилемы. Протофлоэма возникает в кончике корня под перициклом, в межсемядольной плоскости, чередуясь с протоксилемой. На некотором расстоянии от кончика корня, по сторонам от протофлоэмных групп дифференцируются элементы метафлоэмы. Метафлоэмные элементы в дальнейшем продолжают дифференцироваться в центростремительном направлении — навстречу формирующейся от центра корня метаксилеме. Таким образом, уже в корне намечается пучковая структура проводящего аппарата, характерная для всех надземных органов. В гипокотиле пучки полностью обособляются. Последовательность этого процесса можно проследить на серии поперечных срезов от основания корня к основанию семядолей.
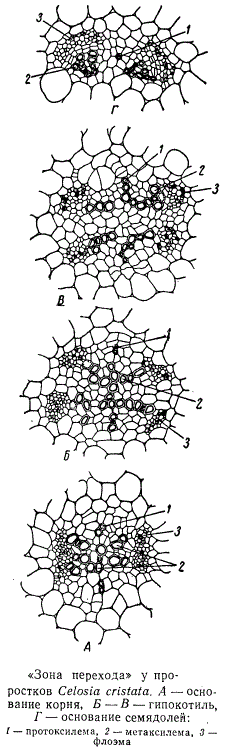
В нижней части гипокотиля, в межсемядольной плоскости, образуется паренхимная прослойка, делящая пополам флоэму, а затем и ксилему. Развитие паренхимной ткани в семядольной плоскости способствует окончательному обособлению коллатеральных пучков, которые по два с каждой стороны тянутся по направлению к семядолям. Между коллатеральными пучками остаются сосуды протоксилемы, окруженные со всех сторон паренхимными клетками. Об изменении положения этих сосудов в зоне перехода уже говорилось. После того как протоксилема займет эндархное положение, паренхимная прослойка между коллатеральными пучками исчезает, образуется единый коллатеральный пучок, составляющий среднюю жилку семядоли.